Тема: Сравнительный анализ жизненных циклов различных представителей Heterophyata (Trematoda: Digenea)
Характеристики работы
Закажите новую по вашим требованиям
Представленный материал является образцом учебного исследования, примером структуры и содержания учебного исследования по заявленной теме. Размещён исключительно в информационных и ознакомительных целях.
Workspay.ru оказывает информационные услуги по сбору, обработке и структурированию материалов в соответствии с требованиями заказчика.
Размещение материала не означает публикацию произведения впервые и не предполагает передачу исключительных авторских прав третьим лицам.
Материал не предназначен для дословной сдачи в образовательные организации и требует самостоятельной переработки с соблюдением законодательства Российской Федерации об авторском праве и принципов академической добросовестности.
Авторские права на исходные материалы принадлежат их законным правообладателям. В случае возникновения вопросов, связанных с размещённым материалом, просим направить обращение через форму обратной связи.
📋 Содержание
ОБЗОР ЛИТЕРАТУРЫ 5
ЦЕЛИ И ЗАДАЧИ 29
МАТЕРИАЛЫ И МЕТОДЫ 29
РЕЗУЛЬТАТЫ 31
ОБСУЖДЕНИЕ 39
ВЫВОДЫ 50
СПИСОК ЛИТЕРАТУРЫ 51
📖 Введение
Существует несколько взглядов на природу жизненного цикла трематод. В первую очередь разногласия возникают в трактовке способа размножения спороцист и редий. Среди некоторых зарубежных исследователей второй половины 19-го, начала 20-го века была распространена точка зрения, согласно которой поколение спороцист-редий возникает за счет особой формы почкования - бесполого размножения (Steenstrup, 1842). Сегодня такую точку зрения нельзя считать актуальной. Почкование всегда захватывает целый пласт клеток. В случае внутримоллюсковых стадий трематод речь идет о единичных клетках, из которых развиваются эмбрионы. Основные споры сосредоточены вокруг вопроса о природе этих клеток. Согласно одной точке зрения (Leukart, 1882; Faust, 1917), которая ближе и автору данной рукописи, это клетки половой линии. Другие авторы, хоть и называют способ размножения спороцист и редий партеногенезом, но расценивают генеративные клетки как элементы сомы (Whitfield, Evans, 1983). В последнем случае говорят опять-таки о бесполом размножении. Также существует взгляд, по которому весь цикл представляет собой растянутый онтогенез одного поколения (Brooks, 1930). Причем, в этом случае, спороцисты- редии возникают вследствие полиэмбрионии. Надо признать, что спор между исследователями, придерживающихся тех или иных взглядов по этому вопросу, несет отчасти семантический характер. Опустив подробности этого спора, можно утверждать, что разногласия в первую очередь говорят об отсутствии ясного понимания судьбы и самой природы структур, входящих в состав тела на той или иной стадии жизненного цикла. Возможно, во многом именно с этим непониманием и связано отсутствие не только самой системы, но и возможности ее создания. В огонь подливает масло то обстоятельство, что для многих групп трематод данные по некоторым стадиям жизненного цикла если и имеются, то весьма фрагментарны. В первую очередь, речь идет о мирацидиях и поколении материнской спороцисты.
Долгое время до начала 20-го столетия никаких существенных попыток создания единой системы трематод не производилось. Различные роды сосальщиков к тому времени только начали объединять в семейства. Такие объединения строились по анатомическим характеристикам половозрелых форм. Первая система трематод, не основывающаяся на морфологии марит, была предложена Люэ (Luhe, 1909). В своей работе автор попытался выделить различные морфотипы церкарий. Забегая вперед, стоит отметить, что, во многом, разделение трематод на две системы (система церкарий и система марит) сохранилось по сию пору. Система Люэ была построена по чертам внешней морфологии церкарий: наличие брюшной присоски, наличие фурки, плавниковой складки, стилет и т. д. В дальнейшем, ряд авторов вводил корректировки и дополнения в систему Люэ (Lebour, 1911; Kobayashi, 1922; Sewell, 1922; Miller, 1926 и др.). В их работах были учтены новые признаки, и их системы включали большее число различных представителей трематод. Фаустом (Faust, 1924) была произведена попытка построить систему, опирающуюся на особенности распределения пламенных клеток в теле церкарии. Дюбуа (Dubois, 1929) были введены термины стеностомные и мезостомый типы церкарий - термины, отражающие характеристии собирательных каналов выделительной системы церкарий. Кульмиацией построений систем трематод по морфологическим признакам церкарий, пожалуй, можно считать работу Ля Рю (La Rue, 1957). В этой системе, включающей большое число различных семейств, все трематоды разделяются на две группы: Epiteliocystidea и Anepiteliocystidea. Признак, заложенный в этих названиях - наличие эпителизированного мочевого пузырька. Более детальные разделения в этой системе согласованы с признаками, использованными в ранних построениях. Работа Ля Рю многократно подвергалась критике. В дальнейшем, на ряде представителей электронно-микроскопическими методами было показано, что стенки мочевых пузырьков, которые Ля Рю подозревал в эпителизованности, оказались синцитиальными конструкциями (Powell, 1972, 1973, 1975 и др.). В настоящий момент нет оснований сомневаться в синцитиальной природе стенок мочевых пузырьков у трематод. Так или иначе, система Ля Рю, пожалуй, самая крупная система, построенная по морфологии церкарий трематод (La Rue, 1957).
Во второй половине двадцатого века многие авторы стали задаваться вопросом о том, какая из стадий (церкария или марита) более пригодна для филогенетических построений. Одни утверждали, что для церкарии как для «личиночной» фазы степень гомоплазии должна быть меньше. Другие пытались опровергнуть аргументы первых. Кэйбл (Cable, 1974) представил филогению 56-ти семейств трематод в виде дендрограммы, построенной по множеству морфологических признаков. Эта дендрограмма опирается в большей степени на морфологию церкарий. Некторые авторы строили деревья, придавая большее значение типу дочерних партенит (Odening 1961). Своего рода разрешением этих споров можно считать попытки нескольких авторов построить комплексную систему, основывающуюся на морфологических характеристиках всех стадий жизненного цикла (Odening, 1974; Brooks et al., 1985). Однако стоит отметить, что в этих построениях по-прежнему большее внимание уделялось признакам церкарий и марит.
С развитием технологий секвенирования нуклеотидных последовательностей к началу 21-го века накопилось большое количество данных по различным группам трематод. Олсон с соавторами (Olson et al., 2003), свели последовательности генов малой и большой субъединиц рибосомы в единые матрицы и построили филодендрограмму, включающую 163 таксона.
Пожалуй, каждая упомянутая реконструкция филогении требует подробного описания и анализа с современной точки зрения. Однако это не входит в задачи данной работы, и, скорее всего, требует некого сотрудничества между исследователями, специализирующимися на разных группах сосальщиков. Не стоит полагать, что дендрограммы, накопленные к сегодняшнему дню малоценны. Если попытаться проследить «эволюцию» реконструкций от системы Люэ до нынешних построений, можно с легкостью заметить, как семейства трематод кластеризуются в монофилетические группировки. Причем эти группировки от системы к системе приобретают большее разрешение (включают большее число семейств). Можно выделить как минимум пять крупных ветвей трематод. Возникают определенные трудности при попытке как-либо их обозначить. Разные авторы предлагают разные названия. Так или иначе, есть основания выделять слудующие ветви: трематоды стригео-шистосомоидного ряда, хемиуридные трематоды, эхиностоматиды, плагиорхииды (sensu Odening, 1974) и описторхииды. Представители каждой ветви обладают рядом апоморфий, указывающих на монофилию этих групп. Основные трудности современной филогенетики трематод заключаются в отсутсвии понимания того, как эти группировки сообщаются между собой. Иными словами, порядок ветвления на существующих дендрограммах оказывается самым разным. В данной работе сосредоточено внимание на трематодах группы Opisthorchiida (sensu Odening, 1974).
Описторхииды - обширная группа трематод, включающая не менее 130-ти родов. Большинство исследователей, занимавшихся этой группой, выделяло три семейства: Cryptogonimidae, Opisthorchiidae и Heterophyidae. Есть серьезные основания сомневаться в монофилии каждого из трех семейств. Поэтому автор данной рукописи остерегается использовать название Opisthorchiida. Вместо него в заглавии и в самом тексте употребляется во многом забытое для этой группы слово - Heterophyata.
Анализируя морфологию отдельных стадий жизненного цикла гетерофиат, можно прийти к выводу, что данная группа обладает множеством апоморфий, которые указывают на эволюционно сложившуюся специализацию этой группы. В то же время некоторые признаки сохраняются здесь в «архаичных» состояниях. В литературе, посвященной трематодам, часто встречается ситуация, при которой авторы ставят представителей гетерофиат в ряд с «примитивными» и в той же работе с «продвинутыми» семействами. Действительно, возникают серьезные трудности при попытке определить место этого таксона среди других ветвей трематод. Таким образом, можно говорить о, своего рода, «промежуточном» состоянии Heterophyata. Накопление новых сведений и попытки выявить базальные для данной группы признаки явились бы возможностью оценить положение гетерофиат в системе трематод. Данная работа является своего рода плацдармом для дальнейших исследований в этой области.
✅ Заключение
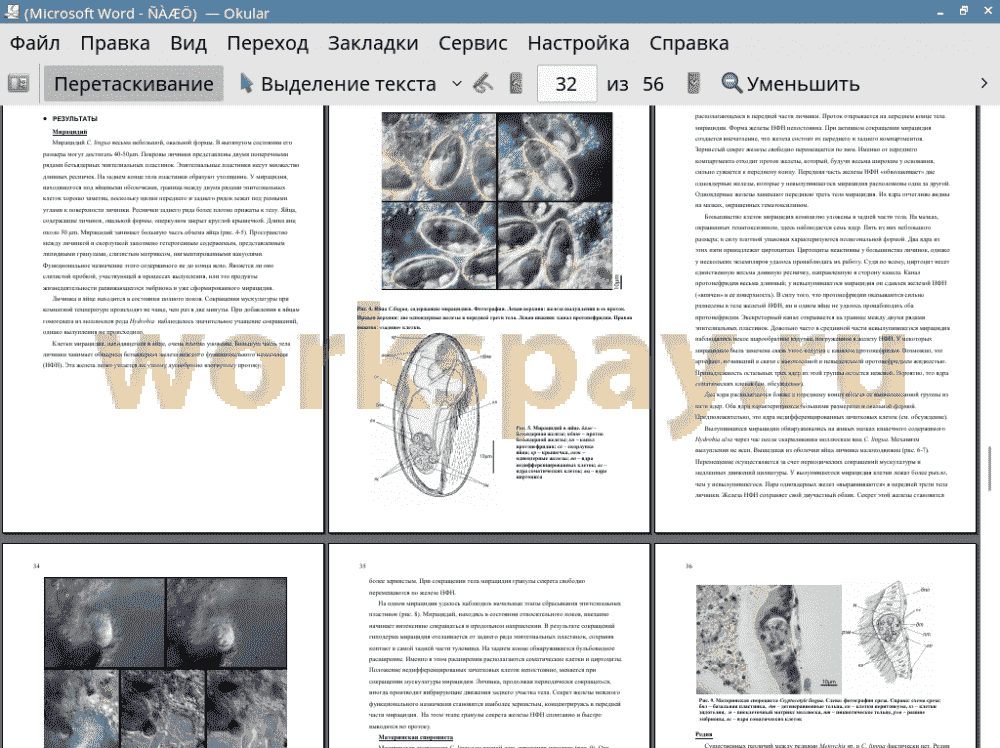
1. Морфология мирацидия Cryptocotyle lingua значительно отличается от морфологии личинок Plagiorchioidea, но особенности железистого аппарата мирацидия C. lingua схожи с таковыми у личинок Microphallidae.
2. Ранние этапы разития материнской спороцисты C. lingua, в целом, схожи с таковыми у Plagiorchiata, однако начало развития эмбрионов дочернего поколения у Cryptocotyle происходит раньше.
3. Редии Heterophyata отвечают единому плану строения. Различия касаются сезонной динамики размножения редий, паразитирующих в морских и пресноводных моллюсках. Редия C. lingua характеризуется типичной погруженной поляризованной герминальной массой.
4. Церкарии Heterophyata характеризуются высокой степенью сходства строения. Различия касаются типа плавниковой мембраны, расположения желез проникновения. Степень различия строения выделительной системы церкарий требует дополнительных исследований.
5. Система Heterophyata построена по характеристикам копулятивного аппарата марит, однако различные варианты строения последнего описываются одним термином (гонотиль). Природа структур, входящих в состав аппарата оплодотворения большинства представителей нуждается в уточнении.
📕 Список литературы
🖼 Скриншоты