Тема: ПРЕОБРАЗОВАНИЯ БИОГЕОЦЕНОЗОВ СУХОДОЛЬНЫХ ЛУГОВ В ПРОЦЕССЕ ЗАРАСТАНИЯ ОЛЬХОЙ СЕРОЙ (ALNUS INCANA (L.) MOENCH)
Закажите новую по вашим требованиям
Представленный материал является образцом учебного исследования, примером структуры и содержания учебного исследования по заявленной теме. Размещён исключительно в информационных и ознакомительных целях.
Workspay.ru оказывает информационные услуги по сбору, обработке и структурированию материалов в соответствии с требованиями заказчика.
Размещение материала не означает публикацию произведения впервые и не предполагает передачу исключительных авторских прав третьим лицам.
Материал не предназначен для дословной сдачи в образовательные организации и требует самостоятельной переработки с соблюдением законодательства Российской Федерации об авторском праве и принципов академической добросовестности.
Авторские права на исходные материалы принадлежат их законным правообладателям. В случае возникновения вопросов, связанных с размещённым материалом, просим направить обращение через форму обратной связи.
📋 Содержание
Введение 2
Материал и методика 3
Результаты и обсуждение 4
Литература 14
References 15
📖 Введение
Одним из видов деревьев, активно участвующих в процессе восстановления лесной растительности на лугах, выступает ольха серая (Alnus incana (L.) Moench). Благодаря особенностям размножения и развития, эта порода быстро заселяет вырубки, заброшенные пашни, сенокосные и выпасные угодья. Важнейшей отличительной характеристикой ольхи является ее способность вступать в симбиотические отношения с азотфиксирующими корневыми актиномицетами (актинобактериями) из р. Frankia (Frankia alni), что позволяет ей в значительной степени улучшать собственное корневое питание и почвенные условия в местах произрастания [14]. Эта порода весьма нетребовательна к условиям среды и способна быстро их модифицировать.
Цель данной работы состоит в изучении поведения ольхи серой на этапах последовательного зарастания суходольного луга. Основные задачи — описать состав и строение сероольховых синузий; исследовать их влияние на изменение условий освещенности, влажности, температурного режима, характеристик почвы и подстилки; оценить влияние на микробиологическую активность почвы; исследовать преобразование состава и биомассы почвенной мезофауны; выявить влияние ольхи на луговую растительность через оценку изменения видового состава и обилия видов напочвенного покрова.
✅ Заключение
1. Зарастание луга ольхой серой ведет к изменению характеристик биотопа: снижению освещенности и значительному увеличению мощности подстилки.
2. В результате зарастания луга ольхой изменяется количественный состав мезофауны верхних горизонтов почвы: значительно снижается численность и биомасса дождевых червей на единицу площади, а таксономическое разнообразие представителей типа Членистоногие в этом направлении увеличивается.
3. Отличий в протеазной активности почвы на участках зарастания ольхи разных возрастов и на лугу не обнаружено: снижение ризосферного эффекта под пологом старой ольхи компенсируется обильной микробиотой, привлеченной богатым азотом листовым опадом.
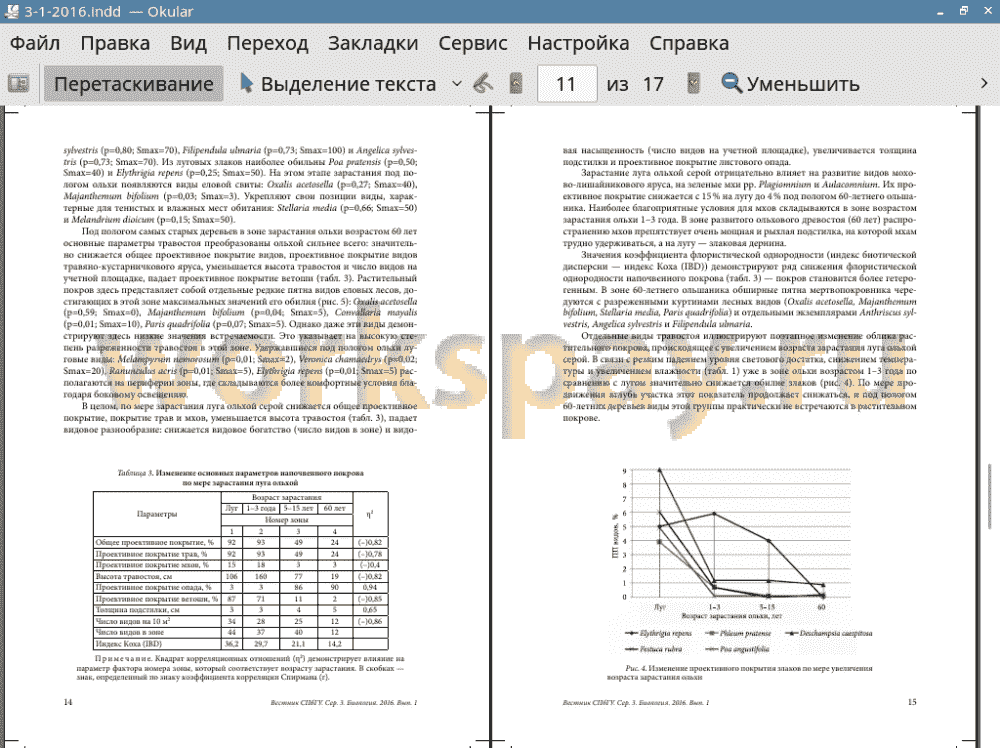
4. Под влиянием ольхи снижается видовое богатство, видовая насыщенность и резко падает обилие (покрытие) травяного покрова.
5. В напочвенном покрове под пологом ольхи значительно снижается участие луговых и возрастает участие лесных видов.
6. Под пологом ольхи по сравнению с лугом возрастает флористическая гетерогенность растительного покрова.
7. При зарастании луга ольхой серой возобновление ольхи благополучно развивается под пологом собственного древостоя, что позволяет ольхе длительно удерживать занимаемую территорию.
8. Отсутствие подроста ели на всех стадиях зарастания ольхой свидетельствует о торможении прохождения автогенной сукцессии, направленной на формирование коренного типа растительности — ельника.
📕 Список литературы
🖼 Скриншоты